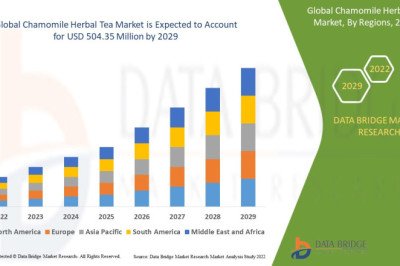
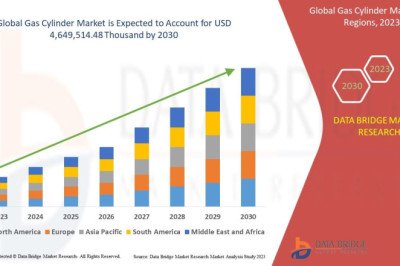
L-alanine is used as a food additive, an ingredient in infusion solutions, and a precursor to chemicals and pharmaceuticals [50]. In Corynebacterium glutamicum, L-alanine is endogenously synthesized from pyruvate through the action of aminotransferases, AlaT and AvtA, where L-glutamic acid and L-valine are the donors of essential amino groups. body molecules [51]. A metabolically engineered Corynebacterium glutamicum strain for L-alanine production was constructed by introducing a gene encoding alanine dehydrogenase (EC 1.4.1.1), which catalyzes the reductive amination of pyruvate [15]. In this biosynthetic pathway, L-glutamate and L-valine are not required for L-alanine synthesis because ammonium ions are the amino donors in the reaction catalyzed by alanine dehydrogenase. To increase alanine production, organic acid production was inhibited by inactivating lactate dehydrogenase and valine structurephosphoenolpyruvate carboxylase [10,15]. The engineered strain produced alanine as the main sugar metabolite under hypoxic conditions, but its productivity was relatively low due to slow glucose consumption [15]. This decrease in glucose consumption is partly due to inefficient reoxidation of NADH leading to an imbalance in reducing equivalents, which leads to impaired glyceraldehyde 3-phosphate dehydrogenase (GAPDH) activity through increased intracellular NADH concentrations [52,53 ]. To increase the rate of glucose consumption under these conditions, the gapA gene encoding GAPDH was overexpressed in the alanine-producing strain, resulting in a marked increase in glucose consumption and alanine production. In addition, elevated activities of 6-phosphoglucose isomerase, 6-phosphofructokinase (PFK) and pyruvate kinase (PYK) also increase glucose consumption and alanine production rates under hypoxic conditions [20]. The performance of this strain was similar to that of the engineered strain GLY3/pCRD500, which overexpresses four genes encoding these glycolytic enzymes, producing 2,430 mM (216 g/L) of alanine after 48 hours of reaction. In general, overexpression of glycolytic genes failed to improve glucose consumption in traditional growth-dependent biological processes [54]. However, this strategy has been successfully used to increase the productivity of L-valine [12], D-lactic acid [19] and isobutanol [21] in addition to alanine in growth-arrested biological processes. A better understanding of the detailed mechanisms involved in improving glucose consumption under hypoxic conditions will lead to ultrahigh productivity of growth-arrested biological processes.
L-alanine is used as a food additive, an ingredient in infusion solutions, and a precursor to chemicals and pharmaceuticals [50]. In Corynebacterium glutamicum, L-alanine is endogenously synthesized from pyruvate through the action of aminotransferases, AlaT and AvtA, where L-glutamic acid and L-valine are the donors of essential amino groups